Duration: 5
min

Yajun Cao
Protein and Antibody Product Marketing
Dr. Yajun Cao graduated from Tsinghua University with a Ph.D. in biology. Yajun
has published her work in journals, including Cancers (Basel) and Stem Cell Research. She joined
GenScript's Antibody Department and has been working as a Senior Scientist since 2024. Yajun has
over 10 years of experience in antibody discovery and antibody-based assay development. She is
currently responsible for mouse monoclonal antibody platform and antibody sequencing platform.
She has participated in the delivery of over 500 mouse mAb project.
The Concept of Anti-Id Antibody
In a specific antibody molecule, the antigenic determinants located in the variable region are
referred to as the idiotope of the antibody. The collective set of all idiotopes and paratopes within an
antibody molecule is termed its idiotype (Figure 1). Antibodies targeting the idiotype of another antibody are
called anti-idiotype antibodies (anti-ID antibodies). The concept of anti-idiotype antibodies was first proposed
by Niels Jerne in 1974 as part of his immune network theory. This theory posits that the immune system functions
as a complex network of interacting antibodies and lymphocytes. It laid the foundation for utilizing
anti-idiotype antibodies as surrogate antigens in vaccines and therapeutics, particularly for targets that are
challenging to produce or tolerate, such as carbohydrates, lipids, or tumor-associated antigens.
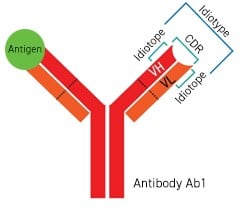
Figure 1: The Idiotope of the Antibody
Classification of Anti-Id Antibody
Anti-idiotype antibodies are primarily classified into four categories based on their binding sites
and functional properties (see Figure 2):
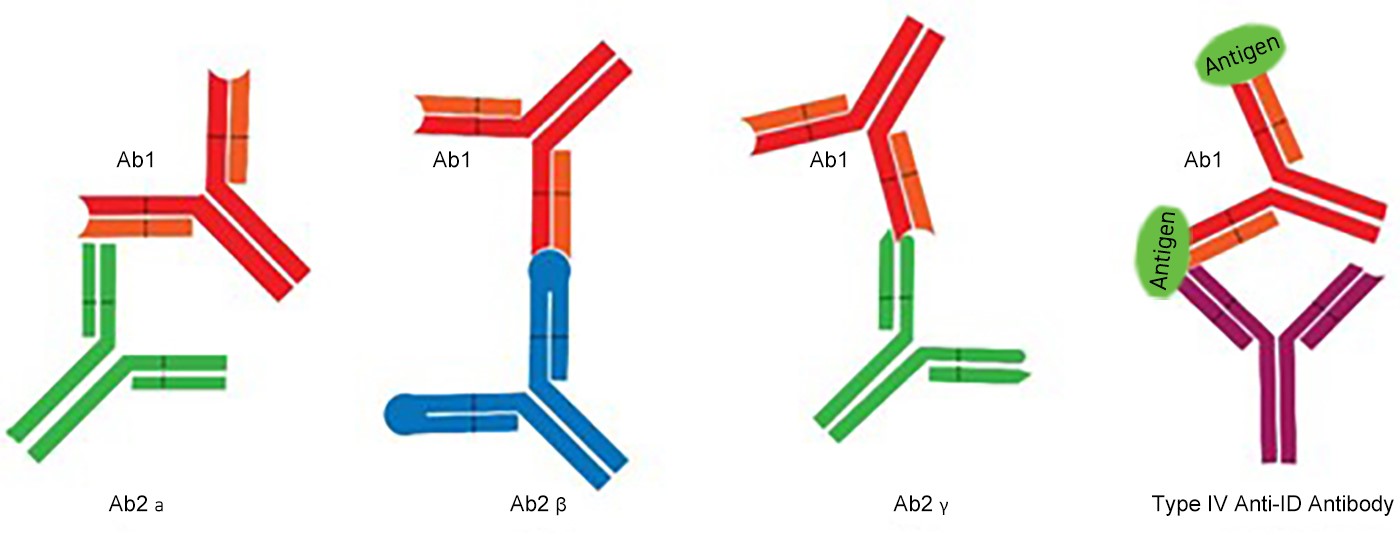
Figure 2: Categories of Anti-ID Antibody
1. Type I Anti-idiotype Antibodies (Ab2α):
These bind to the idiotypic region of Ab1 but not to its paratope (antigen-binding site).
Consequently, Ab2α cannot block the interaction between Ab1 and its antigen, lacking neutralizing or
antigen-binding inhibitory activity.
2. Type II Anti-idiotype Antibodies (Ab2β):
Ab2β specifically recognizes the paratope of Ab1 (the region responsible for antigen binding). By
structurally mimicking the original antigen, Ab2β is termed the "internal image of the antigen". It
competitively inhibits Ab1-antigen binding, demonstrating potent neutralizing and blocking capabilities.
3. Type III Anti-idiotype Antibodies (Ab2γ):
Ab2γ binds to idiotopes adjacent to or partially overlapping with the paratope of Ab1. Unlike Ab2β,
it does not structurally mimic the original antigen. However, it can still interfere with Ab1-antigen
interactions through steric hindrance, thereby exhibiting partial neutralizing activity.
4.Type IV Anti-idiotype Antibodies (Ab2ε):
Generated by immunizing with Ab1-antigen complexes, these antibodies recognize hybrid epitopes
composed of both the idiotope of Ab1 and portions of the bound antigen. Type IV antibodies are particularly
useful for detecting or targeting immune complexes in diagnostic or therapeutic contexts.
Historical Development of Anti-Idiotype Antibody
In the mid-20th century, it was discovered that injecting antibodies into the same or different
species could induce the production of antisera targeting those antibodies. Subsequently, it was found that
antibody molecules possess two types of antigenic determinants: isotypes and allotypes. In 1955, Slater et al.,
while studying the specificity of rabbit antisera against human myeloma proteins, observed that antisera against
certain myeloma proteins, after extensive absorption with unrelated myeloma proteins or normal human
immunoglobulins (Ig), could still react with the original myeloma protein used as the immunogen [1].
They termed
this new antigenic specificity on the antibody molecule as "individual antigenic specificities."
In 1963, Oudin et al. immunized 50 rabbits with Salmonella typhi. They isolated the anti-Salmonella
typhi antibodies from one rabbit's serum and used them as immunogens to immunize normal rabbits [2].
The
resulting antisera contained anti-antibodies. Research revealed that these anti-antibodies could only react with
the anti-Salmonella typhi antibodies used as the immunogen and not with sera from the other 49 Salmonella
typhi-immunized rabbits, nor with sera from rabbits immunized with other antigens or normal rabbit sera. This
indicated that the anti-Salmonella typhi antibodies produced by this particular rabbit possessed unique
antigenic determinants. These determinants differed from those found on antibody molecules produced by other
individuals against the same antigen, as well as from those produced by the same individual against different
antigens. Oudin named these unique antigenic determinants, distinct from isotypes and allotypes, "idiotypes."
In 1974, Jerne, based on modern immune-molecular understanding of antibody idiotypes and building on Burnet's
"clonal selection theory," proposed the famous immune network theory. The immune network theory can be
summarized as follows:
Ab1: The specific antibody produced in response to a foreign antigen, which contains idiotypes
(Id). Ab2: The specific anti-idiotype antibody produced in response to the Id of Ab1. Jerne's immune network
theory emphasized that Ab1 could induce Ab2, and through experiments, he further confirmed the existence of Ab3
[3].
It is well known that the shape of an antibody's antigen-binding region is complementary to the
shape of its antigenic determinant. Similarly, certain Anti-Id antibodies produced against the idiotype of an
antibody have a spatial configuration complementary to the idiotype's antigenic determinant. This double
complementarity results in the antigen-binding region of the Anti-Id resembling the shape and amino acid
distribution of the original antigen's determinant. In other words, from a molecular structure perspective,
Anti-Id with this complementary configuration is equivalent to or approximates the original antigen molecule,
much like a mirror image of the original antigen's determinant. Hence, it is referred to as the "internal image"
of the antigen molecule [2]. In 1991, Nisonoff et al. pointed out that anti-idiotypic antibodies
(specifically
the Ab2β type) could essentially substitute for the antigen in inducing a specific immune response.
Applications of Anti-Id Antibody
Since the discovery of antibody idiotypes in the 1950s, anti-idiotype antibodies have undergone
significant advancements. Initially, these antibodies were primarily utilized for vaccine development (e.g.,
idiotypic vaccines) and the treatment of autoimmune diseases. Today, anti-idiotype antibodies are extensively
applied across multiple domains, including:
- Immunogenicity analysis of antibody-drug conjugates (ADCs),
- Clinical development of anti-drug antibodies (ADAs),
- Controls for ligand neutralization assays and antibody blocking assays,
- Pharmacokinetic (PK) and pharmacodynamic (PD) profiling of antibody therapeutics.
Conclusion
In summary, anti-idiotype antibodies are a fascinating and versatile class of immunoglobulins with
significant potential in both therapeutic and diagnostic applications. Their ability to mimic antigens and
modulate immune responses makes them valuable tools in the fight against a wide range of diseases. However,
realizing their full potential will require continued research and innovation to address the challenges
associated with their production and immunogenicity. As our understanding of the immune system continues to
grow, so too will the opportunities for harnessing the power of anti-idiotype antibodies to improve human
health.
GenScript——One stop Anti-ID antibody development service
Anti-ID antibody development service from GenScript covers diverse types of antigens, including antibody drugs, GCT drugs, peptide drugs, protein drugs, vaccines, oligonucleotide drugs, and small molecule drugs, and provides mouse and rabbit monoclonal antibodies for PK assay development and rabbit-derived ADA for immunogenicity test. The shortest turnaround time for rabbit ADA polyclonal development is 7 weeks, the shortest turnaround time for Anti-ID rabbit monoclonal antibody development is 9 weeks, and the shortest turnaround time for Anti-ID mouse monoclonal antibody development is 10 weeks. GenScript also provides one-stop services such as preparing peptides, proteins, mRNA, and oligonucleotide as antigen. Moreover, both antibody labeling and the development of PK/ADA kits after antibody development can be done at GenScript.
References
[1] SLATER RJ, WARD SM, KUNKEL HG. Immunological relationships among the myeloma proteins. J Exp Med. 1955 Jan 1;101(1):85-108. doi: 10.1084/jem.101.1.85.
[2] OUDIN J, MICHEL M. [A new allotype form of rabbit serum gamma-globulins, apparently associated with antibody function and specificity]. C R Hebd Seances Acad Sci. 1963 Jul 17;257:805-8.
[3] Jerne NK. Towards a network theory of the immune system. Ann Immunol (Paris). 1974 Jan;125C(1-2):373-89.
 Most Popular Reagents
Most Popular Reagents
 Instruments
Instruments
 Antibodies
Antibodies
 Protein Electrophoresis and Blotting
Protein Electrophoresis and Blotting
 Molecular Biology
Molecular Biology
 Stable Cell Lines
Stable Cell Lines
 Cell Isolation and Activation
Cell Isolation and Activation
 IVD Raw Materials
IVD Raw Materials
 Therapy Applications
Therapy Applications
 Resources
Resources